Eight Little Piggies (10 page)
Read Eight Little Piggies Online
Authors: Stephen Jay Gould

History is irrevocable. Once you adopt the ordinary body plan of a reptile, hundreds of options are forever closed, and future possibilities must unfold within the limits of inherited designs. Adaptive latitude is impressive, and natural selection (metaphorically speaking) is nothing if not ingenious. A terrestrial reptile may return to the sea and converge upon fishes in all important aspects of external form. But the similarity can only be, quite literally, skin deep and truly superficial. The convergence must be built with reptilian parts, and this historical signature of an evolutionary past cannot be erased. Dollo explicitly linked his principle of irreversibility with a concept that he called “indestructibility of the past.”
When we look again at the three great convergences of ichthyosaurs—the flippers, the dorsal fin, and the caudal fin—but this time from the alternate perspective of limits imposed by irrevocable starting points, we find that these features beautifully illustrate the three most important principles of irreversibility as a signature of history.
1.
The flippers, or you must use parts available from ancestral contexts little suited to present environments
. The flippers, by external form, are well adapted for swimming and balancing. But their internal bony structure reveals a terrestrial reptile under the marine adaptation. The front flipper begins with a stout humerus, followed by a shortened and flattened radius and ulna, side by side. The carpals and metacarpals (hand bones) and phalanges (finger bones) follow in a similar flattened modification. In an interesting change (still related to an irrevocable ancestral state), the phalanges are multiplied into long rows that mimic the rays of fish fins. Humans have three phalanges per finger (two for the thumb); ichthyosaurs can grow more than twenty per finger.
2.
The dorsal fin, or you can’t get there from here
. The dorsal fin of fishes generally contains a strengthening set of bony rays. Similar structures might well have benefited ichthyosaurs, but their terrestrial ancestors built no recruitable body parts along the back. Ichthyosaurs therefore evolved a boneless dorsal fin (that would have eluded us altogether if we had never discovered the Holzmaden specimens).
3.
The caudal fin and its tailbend, or you must always build a converging structure with some distinctive difference, due to irrevocable ancestry, from the original model
. The vertebral column of fishes, as noted above, either stops at the inception of the tail or extends into the upper lobe. Only in ichthyosaurs do the vertebrae bend down into the lower lobe of the tail. We do not know why ichthyosaurs developed this strikingly different and unique internal structure (I would need another essay to discuss the interesting structural and functional explanations that have been proposed), but all convergences evolve with distinctive differences based on a thousand quirks of disparate ancestries.
Louis Dollo has long been one of my private heros. I meant to cite his views on irreversibility as a centerpiece of this essay, but I didn’t know, until a chance discovery in the midst of my research, that he had written an entire paper on the caudal fin of ichthyosaurs—and at a most interesting time, in 1892 just after the discovery of fin outlines in the Holzmaden specimens. Dollo rejoiced that these beautifully preserved specimens had resolved “
la curieuse dislocation de la colonne vertébrale, signalée depuis longtemps
” (the curious dislocation of the vertebral column, recognized for so long). And he proposed an explanation rooted in uniqueness imposed by irrevocable history. I doubt that he was right in detail, but his conjecture is ingenious, and entirely in the spirit of an important and insufficiently appreciated principle of historical reconstruction. He argued that the tailbend arose because the two-lobed caudal fin of ichthyosaurs evolved from a skin-fold along the back (source of the dorsal fin as well), which extended itself in a posterior direction to form the upper lobe of the tailfin and then pushed the vertebral column down to form the lower lobe. Since several modern reptiles maintain such a skin fold along the back, but never along the belly, new fins could only evolve along the dorsal edge of the body, and the vertebral column could only be pushed down to form a two-lobed tailfin. But ancestral fishes maintained a fin-fold along both back and belly, and a two-lobed tailfin could evolve as a lower lobe pushed the vertebral column up.
Richard Owen, in contrast with his adaptationist colleague Buckland, appreciated the primacy of maintained reptilian design as the main lesson of ichthyosaur convergence. He wrote in his great monograph on British fossil reptiles (published between 1865 and 1881, and anticipating Dollo’s concerns):
The adaptive modification of the Ichthyopterygian skeleton, like those of the Cetacean [whale] relate to their medium of existence; [but] they are superinduced, in the one case upon a Reptilian, in the other upon a Mammalian type.
At about the same time, and in a more pointed commentary on the same theme of irrevocability in history, W. S. Gilbert (in
Princess Ida
) then penned a crisp epitome to remind his audiences of evolution’s major lesson:
Darwinian man though well behaved
At best is only a monkey shaved.
THE MOST SUBLIME
of all beauties often proceed from the softest or the smallest—the quadruple pianissimos of Schubert’s “Schöne Müllerin,” as sung by Fischer-Dieskau (and penetrating with brilliant clarity to the last row of the second balcony, where I once sat for the greatest performance I ever witnessed) or the tiny birds of brilliant plumage depicted in the marginalia of medieval manuscripts. But even the most refined and intellectual character may succumb without shame to the sheer din employed now and then by great composers to overwhelm the emotions by brute force rather than ethereal loveliness—Ravel’s orchestration of the “Great Gate of Kiev” at the end of Moussorgsky’s
Pictures at an Exhibition
, or the last scene of Wagner’s
Die Meistersinger
.
I once had the privilege of singing with the Boston Symphony at Tanglewood in the midst of
numero uno
among musical dins—the
Tuba mirum
of Berlioz’s
Requiem
. I had listened to the piece all my adult life; we had rehearsed (without orchestra) for weeks. I knew exactly what was coming as the dress rehearsal began. The four supplementary brass choirs enter one after the other, building and building to a climax finally joined by the timpani—eight pair, I think, although they seemed to extend forever in an endless row before the choral risers. And against this ultimate crescendo, the basses alone (including me) must sing the great invocation of the last judgment:
Tuba mirum spargens sonum
Per sepulchra regionum
Coget omnes ante thronum
(The wondrous sound of the trumpet goes forth to the tombs of all regions, calling all before the throne.)
So it should go, and so it went—but not for me. I had devolved into tears and spinal shivers—not in ecstasy at the beauty, but in awe at the volume. (Forewarned is forearmed; I was fine at the performance itself.) Great composers have every right to exploit the physiology of emotional response in this way, but only sparingly, for timing is the essence (and most of Berlioz’s
Requiem
is soft).
My memory of this extraordinary incident in my emotional ontogeny focuses upon a curious highlight of mixed modalities. The sound of the brass assaulted my ears, but the thunder of the timpani followed another, unexpected route. It entered the wooden risers under my feet and rose from there to suffuse my body; sound became feeling.
I am no disciple of Jung, and I do not believe in distant phyletic memory. Yet, in an odd and purely analogical sense, I had become a fish for a moment. We (and nearly all terrestrial vertebrates) hear airborne sound through our ears; fish feel the vibrations of waterborne sound through their lateral line organs. Fish, in other words, “hear” by feeling—as I had done through a set of wooden risers with a density closer to water than to air.
For an optimal combination of fascination with excellent documentation, no saga in the history of terrestrial vertebrates can match the evolution of hearing. Two major transitions, seemingly impossible but then elegantly explained, stand out at opposite ends. First, at the inception of terrestrial life: How can creatures switch from feeling vibrations through lateral lines running all over their bodies to hearing sounds through ears? How, in other words, can new organs arise without apparent antecedents? Second, at the last major innovation in vertebrate design: How can bones that articulate the upper and lower jaws of reptiles move into the mammalian ear to become the malleus and incus (hammer and anvil) in the chain of three bones that conduct sound from the eardrum (the tympanum in anatomical parlance, recalling my Berlioz story in the singular) to the inner ear? How, in other words, can organs switch place and function without destroying an animal’s integrity as a working creature? How can we even imagine an intermediary form in such a series? You can’t eat with an unhinged jaw. Creationists have used this difference between reptiles and mammals to proclaim evolution impossible a priori—I mean, really, how can jawbones become ear bones? Get serious! Yet, we shall see, once again, that the domain of conventional thought can be much narrower than the capabilities of nature—although ideas should be able to extend and soar beyond reality.
The key to the riddle of both these transitions lies in the major theme of my Berlioz story—multiple modalities and dual uses. You can pat your head and rub your stomach, walk and chew gum at the same time (most of us, at least), feel and hear sound, chew and sense with the same bones.
Nature writing in the lyrical mode often exalts the apparent perfection and optimality of organic design. Yet, as I frequently argue in these essays, such a position plunges nature into a disabling paradox, historically speaking. If such perfection existed as a norm, you might revel and exult all the more, but for the tiny problem that nature wouldn’t be here (at least in the form of complex organisms) if such optimality usually graced the products of evolution.
I recently made my first trip to Japan to deliver a lecture at the opening of an annual series that will bring one American scholar to Japan and a Japanese counterpart over here to speak on a common topic. I was both pleased and intrigued by our assigned theme for this initial year (largely at Japanese request)—creativity. (Some Japanese apparently fear—although my superficial impressions included nothing to sustain such anxiety—that their scholars and industrialists excel at efficiency and alteration, but not at innovation.)
I had no words of wisdom on Japanese life (I would not dare, not even by the old criterion that experts are folks who have been in a country either more than twenty years or less than two days); nor do I understand the sources of creativity in the human psyche of any culture. So, following the fine maxim that a shoemaker must stick to his last (a wooden model of a foot, not a final goal), I spoke on the evolutionary meaning of creativity—specifically, on the principles that permit major transitions and innovations in the history of life. I don’t know that my message was well received in this land of supreme artistry in the efficient use of limited space, for I held that the watchwords of creativity are sloppiness, poor fit, quirky design, and above all else, redundancy.
Bacteria are marvels of efficiency, simple cells of consummate workmanship, with internal programs, purged of junk and slop, containing single copies of essential genes. But bacteria have been bacteria since life first left a fossil record 3.5 billion years ago—and so shall they probably be until the sun explodes. Such optimality provokes wonder but provides no seeds for substantial change. If each gene does one, and only one, essential thing superbly, how can a new or added function ever arise? Creativity in this sense demands slop and redundancy—a little fat not for trimming but for conversion; a little overemployment so that one supernumerary on the featherbed can be recruited for an added role; the capacity to do several things imperfectly with each part. (Don’t get me wrong. Bacteria represent the world’s greatest success story. They are today and have always been the modal organisms on earth; they cannot be nuked to oblivion and will outlive us all. This time is their time, not the “age of mammals” as our textbooks chauvinistically proclaim. But their price for such success is permanent relegation to a microworld, and they cannot know the joy and pain of consciousness. We live in a universe of trade-offs; complexity and persistence do not work well as partners.)
To build a vertebrate along the tortuous paths of history, evolution must convert the poet’s great metaphor into flesh and bones. “I hear it,” writes Yeats, “in the deep heart’s core.” I don’t mean to be excessively literal, but if creatures couldn’t occasionally hear with their lungs (as some snakes do) or with their jaws (as our immediate reptilian ancestors probably did), we would not now have ears so cleverly wrought that they fool us into the attractive but untenable vision of organisms as objects of optimal design. Consider the first and last major steps in anatomical construction of the mammalian middle ear—for we know no better or more intriguing story in the evolution of vertebrates.
1.
The origin of hearing bones in the first terrestrial vertebrates
. The hearing of sound in thin air poses a major physical problem: How can low-pressure airborne waves be converted into high-pressure waves suitable for transmission by fluids in the cochlea of the inner ear? Terrestrial vertebrates use two major devices to make the necessary conversion. First, on the “stiletto heel” principle (quoting a metaphor from my colleague T. S. Kemp), they collect sound on the relatively large area of the eardrum but eventually transmit the waves into the inner ear through a much smaller opening called the
fenestra ovalis
(oval window). Second, they pass the vibrations from eardrum to oval window along a bone or series of bones, called in mammals, the malleus, incus, and stapes, or hammer, anvil, and stirrup to honor a truly uncanny resemblance. These bones act as levers to increase the pressure as sound waves travel toward the brain.
Fish have an inner ear, but no eardrum or middle-ear bones; they “hear” primarily through their lateral line organs by detecting the movement of water produced by sound waves in this dense medium. How then could middle-ear bones arise in terrestrial vertebrates, apparently from nothing?
The first vertebrates had no jaws. Modern lampreys and hagfishes survive as remnants of this first vertebrate radiation; their formal name, Agnatha (or jawless), embodies their anatomy (or partial lack thereof). In agnathans, a series of gill openings lies behind the boneless mouth—and this arrangement foreshadows the evolution of jaws. In the first jawed fishes, gills are supported by a series of bones, one set for each gill slit. Each set includes an upper and lower bar, pointing forward and hinged in the middle. Obviously, this arrangement, although evolved for supporting gills, looks uncannily like the upper and lower jaws of a typical vertebrate. We do not know for certain whether jaws arose from a functioning gill arch that moved forward to surround the mouth or whether jaws and gill arches just represent two specializations, always separate, but generated from the same system of embryological development. In either case, we do not doubt that gill supports and jaws are homologous structures (that is, evolved from the same source and representing the “same” organ in different forms—like arms and legs or fingers and toes). The evidence for homology is multifarious and overwhelming: (1) the embryo builds both jaw precursors and gill arches not from mesoderm, the source of most bones, but from migrating neural crest cells of the developing head; (2) both structures are made of upper and lower bars, bending forward and hinged in the middle; (3) the muscles that close the jaw are homologues of those that constrict the gill slits.
If vertebrate jaws represent an anterior gill arch, then another crucial element of the skull also derives from the gill supports just behind. The upper bar of the next gill arch in line becomes the hyomandibular of jawed fishes, a bone that functions in support and coordination by linking the jaws to the braincase.
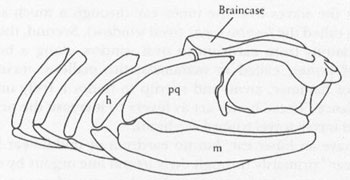
A classic figure of homologies between gill arch and jaw bones, taken from R. L. Carroll’s
Vertebrate Paleontology
and
Evolution
. The upper and lower jaws (pq and m) have the same position and form as all the gill arches behind them. Note also that the upper element of the gill arch just behind the jaws articulates with the braincase. This bone becomes the hyomandibular (h) and later the stapes in terrestrial vertebrates.
Courtesy of Department of Library Services, American Museum of Natural History
.
All this detail may seem distant from the origin of hearing bones, but we are closing in quickly (and shall arrive before the end of this paragraph). Mammals have three middle-ear bones—hammer, anvil, and stirrup, or stapes. And the stapes is the homologue of the hyomandibular in fishes. In other words—but how can it happen?—a bone originating as a gill support must have evolved to brace the jaws against the braincase, and then changed again to function for transmission of sound when water ceded to air, a medium too thin to permit “hearing” by the lateral line.
As usual in a world of encumbrances, we must flush away an old and conventional concept before we can understand how such an “inconceivable” transition might actually occur without impediment in theory or practice. We must forget the old models of horses and humans mounting a chain of improvement in functional continuity—from small, simple, and not-so-good to larger, more elaborate, and beautifully wrought. In these models, brains are always brains and teeth always teeth, but they get better and better at whatever they do. Such schemes may work for the improvement of something already present, for a kind of stately continuity in evolution. But how can something original ever be made? How can organisms move to a truly novel environment, with needs imposed for functions simply absent before? We require a different model for major transitions and innovations, for King Lear was correct in stating that “nothing will come of nothing.”
We need, in other words, a mechanism of recruitment and functional shift. Evolution does not always work by enlarging a rudiment. It must often take a structure functioning perfectly well in one capacity and shift it to another use. The original middle-ear bone, the stapes, evolved by such a route, changing from a stout buttressing bone to a slender hearing bone.