Power, Sex, Suicide: Mitochondria and the Meaning of Life (35 page)
Read Power, Sex, Suicide: Mitochondria and the Meaning of Life Online
Authors: Nick Lane
Tags: #Science, #General

5
Murder or Suicide
The Troubled Birth of the Individual
When cells in the body become worn out or damaged, they die by enforced suicide, or apoptosis. The cell blebs, is packaged up, and reabsorbed. If the mechanisms controlling apoptosis fail, the result is cancer, a conflict of interest between cells and the body as a whole. Apoptosis seems to be necessary for the integrity and cohesion of multicellular individuals, but how did once-independent cells come to accept death for the greater good? Today apoptosis is policed by mitochondria, and the machinery of death was inherited from their bacterial ancestors, suggesting a history of murder. So was the cohesion of the individual forged in deadly conflict?
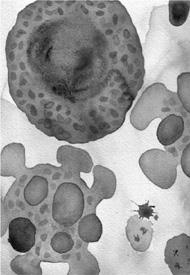
Death by apoptosis—mitochondria determine whether a cell lives or dies by enforced suicide
‘I think therefore I am’ said Descartes, begging the rejoinder ‘But what exactly am I?’ The nature of the individual has long eluded philosophers and scientists, and is only now coming into focus. We can say that an individual is an organism composed of genetically identical cells, which are specialized to perform diverse tasks for the good of the organism as a whole. From an evolutionary point of view, the question is: why did these cells subordinate their selfish interests to collaborate so altruistically in the body? Inevitably there were conflicts between the various levels of organization in the body, between genes, organelles, and cells, but paradoxically without these internecine battles the strong bonds that forge the individual might never have evolved. Such conflicts spurred the evolution of a molecular ‘police force’, which curbs selfish interests much as the legal system enforces acceptable behaviour in society. In the body, programmed cell death,
apoptosis
, is central to the policing of conflicts. Today, apoptosis is enforced by mitochondria, raising the possibility that they may have been key to the evolution of individuals. In this Part we shall see that, back in the mists of evolutionary time, mitochondria were indeed intimately linked with the rise of multicellular individuals.
There has been more spleen vented about selfish genes, altruism, and the limits of natural selection than is seemly in polite scientific society. Underpinning many of the arguments was a simple question: what does natural selection act upon—genes, individuals, groups of individuals (such as a kin group), or the species as a whole? In 1962, Vero Wynne-Edwards’ eloquent treatise on animal behaviour,
Animal Dispersion in Relation to Social Behaviour
, concentrated minds. He ascribed many aspects of social behaviour to selection not at the level of the individual, as had been assumed by Darwin, but at the level of the species. Behaviour was just the tip of the iceberg. Many other traits seemed easier to explain by thinking of the species rather than the individual. For example, ageing doesn’t seem to benefit the individual in any way (what do we gain from getting old and dying?) but does look like a useful service to the species, for it leads to population turnover, preventing overcrowding and over-consumption of lean resources. Similarly, sex seemed pointless for individuals, so much so that we must be bribed by intense erotic pleasure; presumably, mild pleasure is not enough. Rather than simply dividing in two like a bacterium, such that one parent produces two daughter cells, sex takes two parents to produce a single offspring, making it twice as costly as clonal reproduction—the
twofold cost of sex—to say nothing of the trouble of finding a mate. Worse, sex randomizes the very genes that had ensured the success of the parents, making it a potential liability. Its most obvious value is the fast dissemination of variation, and beneficial adaptations, throughout a population: a benefit to the species.
The reaction to these ideas is often dismissed as ultra-Darwinism, a term of disparagement meaning little. How, one must ask, does species-level selection work? There are ways in which it might. For example, a fast population turnover may lead to a fast rate of evolution, which might benefit one species over another if conditions change quickly (for example during rapid global warming, or after a meteorite impact). Another possibility, which Richard Dawkins refers to as the ‘evolution of evolvability’ relates to the genetic ‘flexibility’ of a species—some species have more scope for further evolution in their form and behaviour than others. In most instances, however, the blindness of evolution means that such species-level selection just can’t develop. Sex is complicated and didn’t evolve overnight. If the only benefits are at the species level, and are deferred until sex has actually evolved, what happens in the meantime? Any individuals in a population that take a tentative step towards sex will lose out, and eventually be eliminated by natural selection, because they suffer from the twofold cost of sex and the randomization of any beneficial traits, before any advantages can take over. Similarly, individuals who don’t age will leave their anti-ageing genes behind, which will come to dominate the population simply because the carriers have more time to have more children, who can pass on the same anti-ageing genes. Thus, on one hand there seemed few ways that selection could work at the level of the species, and on the other, some noble self-effacing traits could only be explained (at the time) by recourse to selection at the level of the species.
From the 1960s onwards, William Hamilton, George C. Williams, John Maynard Smith, and others, sought to explain apparently altruistic traits by means of selection at the level of the individual, the kin group, or the gene. The new approach boiled down to a mathematical exploration of
inclusive fitness
—the idea famously expressed in a pub conversation by J. B. S. Haldane: ‘Would I lay down my life to save my brother? No, but I would to save two brothers, four nephews, or eight cousins’ (on the grounds that he shared 50 per cent of his genes with his brothers, 25 per cent with his nephews and 12.5 per cent with his cousins, so his genes at least would break even). Much of the ensuing acrimony centred on the use of such loaded terms as ‘selfish’—terms that have a specific definition in biology, but emotive overtones in general usage. In particular, Richard Dawkins’
The Selfish Gene
either inspired or raised the hackles of an entire generation, at least partly because it was so well written that everyone could feel the icy blast of its conclusion—living organisms are the throwaway
survival machines of their genes, temporary puppets controlled by virtually immortal puppet-master genes. The only logical way to think about evolution, said Dawkins, is to stop gazing at our own belly buttons, and take a genes’-eye view of population dynamics.
The idea that the gene is the ‘unit of selection’ has been attacked from many quarters. The most common line of attack is the claim that genes are invisible to natural selection: they are inert stretches of ticker tape that do no more than code for proteins or RNA. What’s more, there is an ambiguous relationship between a gene and the protein it encodes: the same gene may be split up in different ways, so that it codes for several different proteins; and we now realize that many proteins fulfil more than one function. Genes can also have very different effects, depending on the body they find themselves in. For example, it’s often pointed out that a variant of the haemoglobin gene protects against malaria when present in half dose (heterozygous), but causes sickle cell anaemia when present in full dose (homozygous). All this is true, but none of it undermines the power of a gene-centred approach to explain the currents of evolution: the individual may be the object of selection, but only the genes are passed on to the next generation. The key to the selfish gene is that, in sexual reproduction, the individual does not persist from one generation to the next; no more do any of the individual cells, nor even chromosomes. Bodies dissolve and reform like wisps of cloud, each one fleeting and different. According to Dawkins, only the genes persist, resistant to being scrambled, old as the mountains. From the perspective of a population over evolutionary time, the changes in gene frequencies are the best means of quantifying evolution. To an extent this is a mathematical crutch to a complex problem, but it is also a reality, however unpalatable it may be.
From the point of view of selfish genes, the evolution of an individual is not a problem. If the conglomeration of cells that we call a body happens to be successful at passing on its genes to the next generation, then these genes will thrive to the detriment of the genes that don’t collaborate in this way. A body is the product of genes collaborating together to serve their own selfish end of being copied in ever greater numbers. Dawkins is explicit on the point: ‘Some people use the metaphor of the colony, describing a body as a colony of cells. I prefer to think of the body as a colony of
genes
, and of the cell as a convenient working unit for the chemical industries of the genes.’
The crux of the selfish gene is that only the gene passes from one generation to the next, so the gene is the most stable evolutionary unit: it is the ‘replicator’. Dawkins makes it clear that this perspective is restricted to sexually reproducing organisms, like most (but not all) eukaryotes. It doesn’t apply to bacteria with the same force, because they replicate clonally. In this case, the individual cell
can
be said to persist from one generation to the next, whereas accumulating
mutations mean that the genes themselves
do
change. In fact, in physically stressful circumstances, bacteria can even speed up the mutation rate in their genes. So there is a dilemma in bacteria about whether selection is ‘for’ the genes or the cell as a whole. In many respects the cell is the replicator.
Mutations in a gene don’t necessarily change the phenotype (the function or appearance of the organism) but by definition they must change the gene itself, perhaps even scrambling its sequence out of recognition over aeons. Mutations accumulate because many of them have little or no effect on function, and so go unnoticed by natural selection—they are said to be ‘neutral’. Most of the genetic differences between people, on average one in every 1000 DNA letters, millions of letters in total, are likely to result from neutral mutations. When we consider very different species, two sequences can be so dissimilar that it is not possible to discern any relationship between them, unless we take into consideration the spectrum of intermediary forms in more closely related species. Then we can see that two apparently unrelated genes are indeed related. The physical structure and function of proteins encoded by utterly dissimilar genes is often strikingly well conserved, even though the amino acid components are now mostly different. Plainly, the structure and function of the protein has been selected ‘for’, whereas the sequence of the gene is relatively plastic. It’s like returning to a company that you once worked for, to discover that none of your former colleagues still works there, but that the type of business, ethos, and management structures are exactly as you remembered them, a ghostly echo of the past.
Because genes can change, while the cell and its constituents remain essentially unchanged, the bacterial cell might be considered more stable an evolutionary unit than its genes. For example, cyanobacteria (the bacteria that ‘invented’ photosynthesis) have certainly changed their gene sequences over evolution, but if the fossil evidence can be believed, the phenotype has barely changed over billions of years. If, as Dawkins has argued, the worst enemy of the selfish gene is a competitive (polymorphic, or altered) form of the same gene, then neutral mutations are the selfish gene scrambler
par excellence
: gene sequences diverge over time as neutral mutations accumulate. There may be millions of different forms of the same gene in different species, all scrambled to varying degrees; this is the basis of any gene tree. So evolution pits the selfish interests of genes (which ‘want’ to produce exact copies of themselves) against the randomizing power of mutation, which forever scrambles the sequence of genes, turning the selfish gene into its own worst enemy, the gene it used to hate.