Arrival of the Fittest: Solving Evolution's Greatest Puzzle (20 page)
Read Arrival of the Fittest: Solving Evolution's Greatest Puzzle Online
Authors: Andreas Wagner

In the fall of 2008, I interviewed candidates to conduct a laboratory evolution experiment that could put my nagging doubt to rest. One of them was a young American scientist who stood out, not just because of his credentials but also because he showed up for his interview in hiking boots. While most academic scientists dress informally and would disdain the rigid dress code of other professions, that was still a bit unusual. At least it spoke of a healthy self-confidence.
The young man, Eric Hayden, had just finished his Ph.D. at the University of Oregon, where he had done excellent work on RNA enzymes. A chemist with little prior knowledge about evolutionary biology, he exuded a deep curiosity about innovation. He was instantly likable, thanks to an open face and a smile that lit up the room. After a brief conversation I asked him to chat with other researchers in my group to see whether he would be comfortable in our community. He must have felt right at home, because an hour later he returned to my office in socks—the boots were too warm, he explained.
Eric got the job, and I have never regretted my decision. He was deeply knowledgeable about RNA, a careful experimenter, and a supremely pleasant human being. I felt privileged to work with him.
In my research group, Eric studied a ribozyme, an RNA enzyme that helps some bacteria express their genes. This enzyme recognizes RNA strings with a specific letter sequence, cleaves them, and then attaches one of the string’s fragments to itself.
63
(Many organisms harbor molecules that recognize and cleave specific DNA and RNA texts, for purposes as varied as destroying the foreign DNA of infecting viruses and joining shorter snippets of DNA to form larger meaningful texts.)
64
I had a simple question for this enzyme: Could genotype networks help transform it to recognize a new RNA molecule?
To find out, Eric created more than a billion copies of this enzyme—all of them would fit comfortably into a teaspoon—and used a molecular copy machine to replicate each molecule in this population. This machine is imperfect, because it occasionally makes copying errors, thus sprinkling the population’s ribozymes with mutations. Eric then used a chemical trick to let only some of those mutants replicate—those that could still react with their RNA target. The trick satisfied natural selection’s key requirement that a molecule’s function be preserved for its survival to the next generation.
Eric’s experiment cycled multiple times—think of each cycle as a generation—between this error-prone replication and selection. Before the first cycle, all the molecules in his population were identical, like a billion readers hunched over the same volume in a library. After the first generation, many molecules had already mutated, and only some mutants had survived. The survivors mutated further in the second generation, and so on, until after a mere ten generations, the molecules in the population differed from the starting RNA by an average of five letters, and some by as many as ten letters. The billion readers had spread out in the library.
This simple observation concluded a first part of Eric’s experiment. The population now contained many different RNA molecules, all connected to their parents through a series of single-letter changes. Forced by selection, the molecules in the population had preserved their phenotype, even though their genotype had changed. Because the population had spread through the library, Eric’s experiment had shown that a genotype network exists for RNA enzymes with this phenotype.
The second part of the experiment involved two populations. The first population was the one I just described, while the second was like the starting population, with all identical molecules. Eric supplied both populations with a new and different RNA string to cleave. In this string he had replaced one atom of phosphorus with an atom of sulfur, which makes the enzyme’s job much more difficult. He then evolved both populations separately, through multiple rounds of replication and selection, except that now he selected only molecules that could cleave the
new
RNA string as well. He then asked which populations got the hang of this new task more quickly, the first, spread out over a genotype network, or the second, concentrated in one spot.
If genotype networks help innovation, then the first population should do better, because its members can explore more neighborhoods in the RNA library. And this is exactly what Eric found. The spread-out population found an RNA molecule that excelled at the new task eight times more rapidly than the highly concentrated population.
65
Eric’s experiment also harbored another surprise. This one occurred when we read the letter sequence of the best new molecule that evolution had found, the molecule that was superior to all others at the new job.
Many researchers have studied the RNA enzyme with which we started our experiment. It is a short molecule with only some two hundred letters. We know its letter sequence, its fold, what it does, and how it does it. We know most of what you might want to know about it. And we precisely controlled the environment in which the enzyme evolved, down to the concentration of every molecule, for many generations. With this near-perfect knowledge, you might think that we should be able to
predict
how the molecule would change to solve its new task. If you knew a machine inside out, every single cog, bolt, lever, and spring, and how they work together, you would surely know how best to improve the machine.
We didn’t. The solution that nature came up with to improve our enzyme was completely unexpected. To this date, we do not fully understand why it works best.
Such surprises occur time and again in laboratory evolution experiments. No matter how well a molecule is studied, no matter how simple the experiment, no matter how precisely controlled, nature never ceases to surprise. Even simple enzymes are more difficult to understand than most human machines.
But although we failed to predict a specific solution, we had managed to predict something more important: that a genotype network could accelerate the population’s discovery of this solution. And this prediction was right on target. We can predict innovability even if we cannot predict individual innovations.
Many nonscientists feel troubled when science demystifies nature, identifies the laws that rule it, and thus takes away from their feelings of wonder and awe about the world. In the words of the poet John Keats, a scientist is the sort of killjoy who would “clip an Angel’s wings” and “unweave a rainbow.”
66
That sentiment surely was one reason why Darwin’s theory met resistance, but an experiment like this shows us that we can have it both ways.
67
Science can explain
general
principles of innovability even if it cannot predict any
individual
innovation. Understanding innovability can leave the magic of innovation intact. And that, by itself, is reason for wonder and awe.
CHAPTER FIVE
Command and Control
I
t’s hard to beat milk as a metaphor for goodness. Lady Macbeth’s husband is too full of the “milk of human kindness” to commit regicide, the third chapter of Exodus promises the Hebrews a “land flowing with milk and honey,” and to this day we call harmless things “safe as mother’s milk.” But for more than half of the world’s population, a healthy glass of milk is decidedly not good. It means bloating, gas, and diarrhea. The reason is a lack of lactase, the enzyme that prechews the milk-sweetening sugar lactose for our bodies. Without it, bodies cannot break down lactose, leaving it to gut bacteria that happily scavenge this unused fuel and leave waste products with nasty side effects.
When those lactose-intolerant adults were babies, they could digest the sugar in their mother’s milk just fine. Their lactase gene was turned on—in technical terms, it was
expressed
—which means that their bodies transcribed the DNA instructions for lactase into RNA and translated this RNA into the needed enzyme. The bodies of lactose-intolerant adults have switched the lactase gene off permanently and no longer express it. Genes like the lactase gene, which our bodies can turn on or off, are
regulated
genes
.
For most of human history, the “off” position in adults was the norm. If you are lucky enough to tolerate lactose, you have a mutation in the lactase control region, a stretch of DNA near the lactase gene that leaves the lactase gene turned on well into adulthood. Chances are that your distant ancestors were milk-drinking cattle farmers, because mutations that cause lactose tolerance first spread through pastoral populations, like those of East Africa and Scandinavia. And they spread blazingly fast, from zero to more than 90 percent of some populations, in a blip of time, the eight thousand or so years since humans first discovered the pastoral lifestyle. They are among the strongest recent signatures that natural selection has left in our genomes.
1
Surprising as it may seem, lactose-induced indigestion is deeply connected with innovation. What connects them is regulation—the tuning of the activity of molecules like the lactase gene. Accounting for much more than intestinal upset, regulation is also complicit in the endlessly varying forms of organisms, the gracefully undulating umbrella of a jellyfish, the lethal torpedo of a shark’s body, the slender stem of a rose, the gargantuan trunk of a redwood tree, the deadly coil of a viper, the light-footed legs of a rabbit, and the soaring wings of a bird. Regulation has come a long way from its murky origins in the first cells, where it balanced the growth of a membrane container with that of an RNA genome. More than three billion years later, regulation is shaping the bodies of every living thing on the planet. And no understanding of innovability would be complete without grasping how new regulation appears.
Although regulation controls the form and function of even the most complex organism, like so much else it is most easily studied in the simplest cells, those of bacteria. This is how two French geneticists, François Jacob and Jacques Monod, won their Nobel Prize. Starting in the 1950s, at a time when the double helix had just been discovered, they showed how primitive bacteria like
E. coli
regulate the expression of the genes that permit them to digest lactose.
2
Gene expression begins with the kind of molecular copying machine that we briefly encountered in Eric Hayden’s experiment in chapter 4. It is a
polymerase
enzyme that makes—the name says it all—a polymer, a stringlike molecule consisting of many smaller building blocks, the four nucleotides that we find in the faithful RNA transcript of a gene.
3
When this polymerase transcribes a gene, it first attaches to the gene’s DNA, slides along this DNA letter by letter, and strings together an RNA molecule whose letter sequence is identical to that of the gene.
4
This is also how bacteria express the gene for
their
variant of lactase, an enzyme called beta-galactosidase.
5
(The name is cumbersome, hence it is often abbreviated as beta-gal.) This enzyme cleaves lactose into the two simpler sugars glucose and galactose, from which other metabolic enzymes can extract energy and carbon.
To regulate the beta-gal gene, cells manipulate its transcription with a
transcriptional regulator
. This protein does mostly one thing: It latches on to short stretches of DNA near a gene. Inside the liquid environment of a cell, multiple kinds of regulators drift this way and that, and whenever any one of them encounters a specific DNA sequence—a DNA “word”—it will bind and stick to it. Different regulators have different keywords—the beta-gal regulator recognizes one that contains the letters GAATTGTGAGC.
6
What enables this recognition is the same folded protein shape that makes enzymes work. Regulator and DNA need to have complementary shapes, a bit like Lego blocks where several small studs on one block fit snugly into indentations on another. The analogy is apt but also limited, because shape is not all that matters. For instance, the two molecules also need to have complementary charges or they may repel each other. And where the standard set of Lego blocks has only a few dozen shapes, molecules have many more, tens of thousands in proteins and even more in DNA, where there are as many shapes as there are possible words.
7
What is more, unlike Lego blocks, many molecules spontaneously change shape, not only when they vibrate like enzymes but also when they bind one another. This shape change is similar to what happens when you insert the right key in a lock: Only then can the lock’s cylinder turn and open a door—although in molecules nothing but heat is doing the turning.
The regulator’s Lego-like binding modulates beta-gal production in the simplest possible way: It creates a roadblock for the polymerase, because the regulator’s keyword is placed right where the polymerase starts to transcribe (upper part of figure 13).
8
When no lactose is around, the regulator (R) binds this word and blocks the polymerase (P) from reading the gene—the gene remains turned off.
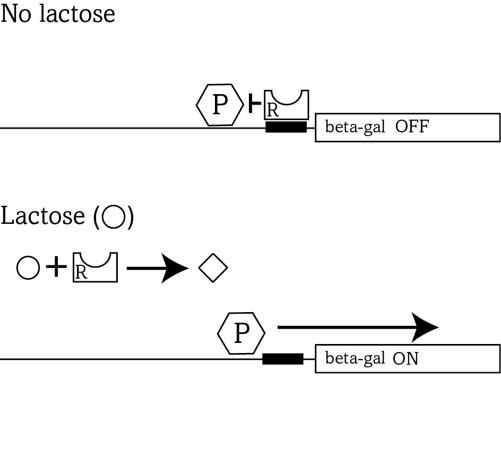
FIGURE 13.
Gene regulation
To use lactose, cells need to get rid of this roadblock whenever the energy-rich substance appears. To understand how they do that, it helps to know that the regulator can bind not only DNA, but also another molecule—like one Lego block that can connect to several others. The other molecule is lactose itself. And when the key of lactose binds the lock of the regulator, the regulator changes shape (see the diamond shape in figure 13) and is no longer complementary to DNA (lower part of figure 13). It detaches from the DNA, which the polymerase can now transcribe freely, letter by letter, into the RNA from which the cell will manufacture the beta-gal protein. In sum, the beta-gal gene is on—beta-gal is produced—whenever lactose is available, but otherwise the gene is off, because its transcription is blocked.
Beta-gal is great stuff. But it isn’t cheap. A cell that expresses beta-gal doesn’t just contain a few dozen of the beta-gal proteins, but rather some three thousand identical molecules, each of them comprising more than a thousand amino acids that need to be manufactured and strung together, which has to be paid for in molecular materials and energy.
9
Common sense suggests that cells should regulate beta-gal to avoid wasting these materials, but if common sense were the surest guide to nature’s ways, biologists would have little to do. The cost to produce beta-gal constantly may well be negligible, given the millions of other molecules that cells manufacture. And leaving the gene on all the time could have a real advantage too, a head start when lactose becomes available.
In 2005, Erez Dekel and Uri Alon from the Weizmann Institute in Israel wanted to find the true cost of expressing beta-gal.
10
They tricked cells into believing that lactose was around when in fact it was not. The cells turned on the beta-gal gene without reason, and if that wastefulness makes a difference, they should divide more slowly. And indeed they did, by a few percent. It’s a bit as if a cash-strapped developer built a house with an unnecessary swimming pool, which cost him money and materials that should have been used to build other, essential rooms. A better builder would finish faster, sell the house, and start a new one, while the other one is still agonizing over the tiles to choose for the pool.
A construction delay amounting to only a few percent may not seem like much, a single minute or so, added to the twenty minutes that
E. coli
cells need to divide. But that minute will eventually be deadly. A population that initially contains 50 percent of such wasteful cells would contain less than 1 percent after eighty days, and fewer than one in a million after merely three hundred days. Rapidly, inevitably, and fatally, they get washed out by the faster-reproducing cells. This is natural selection in rapid and brutal action.
If regulation matters because it avoids waste, then it should be everywhere. And indeed it is. Think of a metabolism with its hundreds of reactions—lactase catalyzing only one of them—as a sophisticated interconnected network of pipelines. Into this network flow nutrients, out of it flow biomass molecules. Each pipe has a dedicated pump, an enzyme that propels materials through it. A cell can regulate each pump according to its needs. If new nutrients turn up in a patch of soil—a fallen apple, a rotting carcass—the soil bacteria turn up the pumps through which these molecules flow. Once the nutrients are gobbled up, they shut these pumps down. And if more of some nutrients and less of others become available, cells can fine-tune the pumps to the right speed.
The beta-gal gene is
repressed
by a regulator, but other genes are regulated in the opposite way: Cells leave them off by default and
activate
them only when needed—through transcriptional regulators that help rather than prevent DNA polymerase from transcribing a gene.
11
And even though the regulation of transcription is the most important kind of regulation, there are many others. Cells regulate how fast they manufacture proteins from transcribed RNA, how active these proteins are, how long-lived, and on and on. This is perhaps the most convincing evidence for regulation’s importance: Life has invented a dozen different ways of doing it.

Imagine the kitchen of a high-end restaurant. The pantry is well stocked with all manner of vegetables, meats, fruit, fish, cooking oils, spices, and flavorings. There are enough ingredients to create every conceivable dish, from the mundane to the exotic, each one nutritious and delicious. The executive chef wants a complete supply of everything, all the time. One of regulation’s roles in metabolism is that of a penny-pinching manager, requisitioning just the right amount of ingredients, anxious not to squander money on even as little as an extra potato.
But regulation is much more than that. It also provides the recipes, the kind that call for cooking a cup of beans with two cups of chicken stock and a dash of salt in a 350-degree oven for thirty minutes. Each recipe is a sophisticated gene expression program encoded in the genome. It tells cells just when and how much to make of each protein ingredient in an organism.
If comparing a blue whale to a soufflé seems offensive, consider the complexity of life’s recipes. Each cell type needs many more ingredients than even the most complex dish, thousands of them, in amounts so finely tuned and so exquisitely timed that not even the most skilled five-star cook could hope to follow life’s recipes. What is more, evolution has untiringly created new recipes that brought forth ever-new dishes, innovations in cells, tissues, and organs, as well as entirely new
kinds
of bodies that emerge through the shifting and enormously complex patterns of regulation.