Eight Little Piggies (35 page)
Read Eight Little Piggies Online
Authors: Stephen Jay Gould

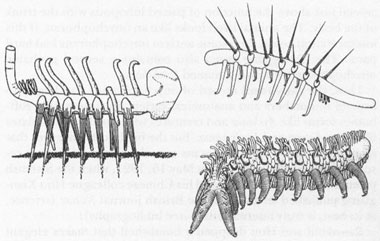
Left: Conway Morris’s original reconstruction of
Hallucigenia. Simon Conway Morris (1977): Reprinted by permission of
Palaeontology. Right top: Ramsköld and Hou’s inversion of
Hallucigenia
as an onychophoran.
Reprinted by permission from
Nature; Copyright © 1991 Macmillan Magazines Limited. Right bottom: Ramsköld and Hou’s reconstruction of the new Chengjiang onychophoran with side plates and spines.
Reprinted by permission from
Nature;
Copyright © 1991, Macmillan Magazines Limited
.
Hallucigenia
is so peculiar, so hard to imagine as an efficiently working beast that we must entertain the possibility of a very different solution. Perhaps
Hallucigenia
is not a complete animal, but a complex appendage of a larger creature, still undiscovered. The “head” end of
Hallucigenia
is no more than an incoherent blob in all known fossils. Perhaps it is no head at all, but a point of fracture, where an appendage (called
Hallucigenia
) broke off from a larger main body.
I received several dozen much appreciated letters from readers of my book, suggesting different reconstructions for some of the oddball creatures of the Burgess Shale.
Hallucigenia
received the lion’s share of attention—and one suggestion cropped up again and again, in at least twenty separate letters. These correspondents, nearly all amateurs in natural history, pointed out that
Hallucigenia
would make much more sense turned upside down—for the spines, which never made any sense as organs of locomotion, could then function far more reasonably for protection!
I responded to these letters with, for me, the decisive rejoinder that a single row of tentacles (Simon’s version of the upper surface) would work even more poorly than paired spines as devices of locomotion. How could an animal balance, not to mention hop around, on a single row of tentacles? Yet I couldn’t deny that everything else made more sense upside down.
It doesn’t happen often, but if Ramsköld and Hou are correct—and I think they are—then the gut feeling of amateurs has triumphed over the weight of professional opinion. For Ramsköld and Hou have, unbeknownst to them of course, followed the advice of my correspondents. They have turned
Hallucigenia
upside down, but with an added twist (intellectual, not geometric) as well—they have inverted it into an onychophoran!
Ramsköld and Hou present two major arguments for their inversion of
Hallucigenia
. First, they must tackle the issue that hung me up: How can a single row of tentacles function as legs? They acknowledge the problem, of course, but suggest that Conway Morris was wrong and that two rows of paired tentacles are actually present along the surface that he called dorsal, or topmost. If Ramsköld and Hou are correct, then the major objection to reversing
Hallucigenia
disappears—for two rows of flexible tentacles look like the ordinary legs of a bilaterally symmetrical creature. Moreover, when you turn
Hallucigenia
upside down on the assumption that two rows of tentacles adorn the topside of Conway Morris’s version, then the inverted beast immediately says “onychophoran” to any expert, for the little paired pincers at the end of each tentacle become dead ringers for onychophoran claws. Ramsköld and Hou have not yet developed enough evidence to prove the double row of tentacles conclusively, but our museum at Harvard contains the sample best suited for resolving this issue—a slab of rock with more than a dozen
Hallucigenia
specimens. I have lent this slab to Ramsköld and Hou, and I suspect that an answer will soon be forthcoming.
Second, they must explain how an onychophoran could possess the several pairs of long, pointed, upward-protruding spines that an inversion of
Hallucigenia
places along the top edge of the animal—for some fossil onychophorans bear plates. (
Microdictyon
and
Luolishania
as previously discussed), but none yet described carry spines. Here, Ramsköld and Hou present compelling evidence in a form much favored by natural historians—a sequence or continuum linking a strange and unexpected form to something familiar through a series of intermediates.
Start the series with
Microdictyon
. This animal, probably an onychophoran, carried pairs of flat plates along the side of its body just above the insertion of lobopods. Go next to a new and as yet unnamed “armored lobopod,” again from the prolific Chengjiang fauna. This clear onychophoran also bore paired plates in the same position as in
Microdictyon
. But each plate now carries a small spine (see figure)—nothing like the elongation in
Hallucigenia
, but evidence that onychophoran plates can support spines. For a third step, go to isolated plates collected in lower Cambrian rocks of North Greenland by J. S. Peel and illustrated by Swedish paleontologist Stefan Bengtson in a commentary in
Nature
written to accompany Ramsköld and Hou’s paper. These Greenland plates have the same meshwork structure as those of
Microdictyon
—and onychophoran affinity seems a reasonable conjecture (although in this case, we have only found preserved plates, not the entire body). But the Greenland plates carry spines verging on the length of
Hallucigenia
spikes. We now require only a small step to a fourth term in the series—to an onychophoran bearing plates with highly elongated spines: in other words, to
Hallucigenia
turned upside down!
We are witnessing a veritable explosion of Cambrian onychophorans—
Aysheaia
and
Xenusion
with their soft bodies,
Microdictyon
and
Luolishania
with plates, the unnamed Chengjiang creature with plates and short spines, the Greenland form with longer spines, and finally, inverted
Hallucigenia
with greatly extended spines.
The reversal of
Hallucigenia
has capped and sealed the tale. The larger conclusion seems inescapable: In the great period of maximal anatomical variety and experimentation that followed right after the Cambrian explosion first populated the earth with multicellular animals of modern design, the Onychophora represented a substantial and independent group of diverse and successful marine organisms. The modern terrestrial species are a tiny and peripheral remnant, a bare clinging to life for a lineage that once ranked among the major players. The shoehorn and straightening rod have served us poorly as strategies of interpretation. Groups with few species may be highly distinct in genealogy.
Onychophorans, moreover, are not the only small cluster of straggling survivors within groups that were once major branches of life’s tree. The distinct phylum of priapulid worms, for example, contains fewer than 20 species worldwide, compared with some 8,000 for marine polychaete worms, members of the dominant phylum Annelida. Yet, in the Cambrian period at the beginning of multicellular history, priapulids and polychaetes were equally common and similarly endowed (or so it seemed) with prospects for long-term success. Moreover, just as onychophorans have held on by surviving in the peripheral habitat of terrestrial life (for a formerly marine group), modern priapulids all live in harsh and marginal environments—mostly in cold or deep waters and often with low levels of oxygen.
In recognizing the Onychophora as a distinct group with an ancient legacy of much greater breadth, we may regret the loss of tidiness provided by the shoehorn and straightening rod, but we should rejoice in the interesting conceptual gains. For by our latest reckoning of life’s early history, “uncomfortable” groups like the Onychophora should exist today. We once thought that the history of life moved upward and outward from simple beginnings in a few primitive, ancestral lines to ever more and ever better—the conventional notion that I have called the cone of increasing diversity. On this model, an ancient and distinctive genealogical status for several small groups (like the Onychophora) makes no sense—for life’s early history, at the point of the cone, shouldn’t have featured many distinct anatomies at all. The large living groups of mollusks, arthropods, annelids, vertebrates, etc.—all of which have fossil records extending back to this beginning—provide quite enough material for legacies from these early times of limited simplicity. But the reinterpretation of the Burgess Shale, and our burgeoning interest in the early history of multicellular life in general, have indicated that the cone model is not only wrong but also backward. Life may have reached a maximal spread of anatomical experimentation in these early days—and later history may be epitomized as a diminution of initial possibilities by decimation, rather than a continual expansion.
In this reversed model of a grass field, with most blades clipped off and just a few proliferating wildly thereafter, we should expect to find some blades that survived the mower but never flowered extensively again—whereas, in the cone model, the forest of blades never existed, and the early history of life provides insufficient raw material for many distinct modern groups like the Onychophora.
However much I may regret the loss of a wonderful weirdo in the reversal of
Hallucigenia
, and in its consequent change in status from oddball to onychophoran, I am more than compensated by fascinating insight into the history of ancestry for my favorite name bearer,
Peripatus
. I revel in the knowledge that these marginal and neglected animals belong to a once-mighty group that included armored members with plates and long spines. And I rejoice in the further knowledge thus provided about the strange and potent times of life’s early multicellular history. (My regret, in any case, could not possibly be more irrelevant to nature’s constitution, either now or 500 million years ago.
Hallucigenia
was what it was. My hopes, and those of any scientist, are only worth considering as potential biases that can block our understanding of nature’s factuality.)
Peripatus
may walk prouder in the pleasures of pedigree. We humans, as intellectual descendants of Aristotle, the original peripatetic, might consider a favorite motto from “the master of them that know”—well begun is half done (from the
Politics
, book 5, chapter 4). Apply it first to the onychophorans themselves—for in a tough world dominated by contingent good fortune in surviving extinction, a strong beginning of high diversity affords maximal prospect for some legacy long down the hard road. But apply it also to us, the paleontologists who strive to understand this complex history of life. By turning
Hallucigenia
upside down, we have probably taken a large step toward getting the history of life right side up.
TWO SUCCESSIVE
symbols of Saint Louis typify the passages of our century. Saarinen’s magnificent arch, gleaming and immaculate, seems to soar from the Mississippi River into heaven (an optical illusion, in large part, cleverly produced by a gradual decrease in edge length from 54 feet at the base of the structure to 17 feet at the summit. Our minds expect a constant size, and the marked decrease therefore makes the summit seem ever so much higher than its actual 630 feet—the size of an ordinary skyscraper in a modern city). By contrast, St. Louis’s older symbol, an equestrian statue of the eponymous Louis IX, the only canonized king of France, still stands in front of the art museum in Forest Park. It is anything but immaculate, thanks to that primary spotting agent of all cities—pigeons.
As a team, pigeons and the statue of Saint Louis go way back. The current statue is a 1906 recasting in bronze of the impermanent original made for the main entrance to one of the world’s greatest expositions—the 1904 World’s Fair held to celebrate (if just a bit late) the centenary of the Louisiana Purchase. The fair must have been spectacular; my wife’s family, raised in Saint Louis, still mentions it with awe in stories passed down through three generations. The fair gave us iced tea, ice cream cones, and a great song, “Meet Me in Saint Louis, Louis” (many folks don’t even know the next line—“meet me at the fair”).
A ferris wheel stood twenty-five stories high; Scott Joplin played his rags. The Pike, main street of the amusement area, featured daily reenactments of the Boer War and the Galveston Flood. The world’s greatest athletes came to participate in the third Olympic Games. The fairgrounds, bathed at night in the newfangled invention of electric lighting, inspired Henry Adams to write: “The world has never witnessed so marvelous a phantasm; by night Arabia’s crimson sands had never returned a glow half so astonishing [a statement that will need revision after the night bombing of Desert Storm], as one wandered among long lines of white palaces, exquisitely lighted by thousands and thousands of electric candles, soft, rich, shadowy, palpable in their sensuous depths” (from
The Education of Henry Adams
). This statement also makes sense of the next two lines of the famous song: “Don’t tell me the lights are shining/Any place but there.”
Intellectuals must be constantly clever and industrious. We know that we are peripheral to society’s main thrust, and we must be constantly vigilant in seeking opportunities to piggyback on larger enterprises—to find something so big and so expensive that prevailing powers will grant us a bit of space and attention at the edges. The hoopla and funding of major exhibitions often gives us a little room for a smaller celebration in our own style. I was invited to give a speech at something called the “Academic Olympiad” in Seoul during the last Olympic games. (I wasn’t able to go and never heard boo about the outcome—though television deluged us with details about javelins and the hop, step, and jump.) Similarly, since the 1904 World’s Fair set up shop right next to Washington University, academicians rallied to hold a “Congress of Arts and Science” at the Universal Exposition (as the fair was officially called). At this collocation, the great American biologist Charles Otis Whitman gave a leading address with the general title: “The Problem of the Origin of Species.” He spoke primarily about pigeons.
Whitman’s work, while treating so humble a subject, had a certain panache and boldness. He wrote at a time when biologists, though fully confident about the fact of evolution, had become very confused and polemical about the causes of evolutionary change. Darwin’s own theory of natural selection had never commanded majority support (and would not emerge as a general consensus until the 1930s). As visitors ate their ice-cream cones on the Pike, at least three other theories of evolutionary change enjoyed strong support among biologists—(1) the inheritance of acquired characters, or
Lamarckism
; (2) the origin of species in sudden jumps of genetic change, or
mutationism
; (3) the unfolding of evolution along limited pathways set by the genetic and developmental programs of organisms, or
orthogenesis
(literally, “straight line generation”). Whitman, who had been raising and breeding pigeons for decades, wrote his article to defend the last alternative of orthogenesis, thereby relegating Darwinian natural selection to a small and subsidiary role in evolution.
Whitman’s boldness did not lie in his choice of the orthogenetic theory—for this argument was a strong contender in his day, though probably the least popular of the three major alternatives to Darwinism. We judge a man intrepid when he uses his adversary’s tools or data to support a rival system. A famous story about Ty Cobb tells of his disgust with Babe Ruth’s new style of power hitting (Ruth swatted more home runs per year all by himself than most entire teams had formerly managed in a season). Cobb, the greatest and most artful practitioner of the earlier style of slap, hit, and scramble for a run at a time, held his hands apart, slipped them together high on the bat as the pitch came in (thus achieving maximal control while sacrificing power), and then slapped the ball to his chosen spot; Ruth, by contrast (and following the strategy of all sluggers), held the bat at the end and swung away, missing far more often than he connected. Cobb regarded this style as easy and vulgar, however effective. One day, near the end of his career, and to show his contempt in the most public way, Cobb ostentatiously held the bat in Ruth’s manner, hit three home runs in a single game and then went right back to his older, favored style forever after.
Whitman’s assault on Darwin’s theory from within was far bolder and more sustained, if not quite so showy. For Whitman had chosen, for study over decades, the very organisms that Darwin had selected as the primary empirical support for his own theory—pigeons.
Darwin stated in chapter 1 of the
Origin of Species
:
Believing that it is always best to study some special group, I have, after deliberation, taken up domestic pigeons. I have kept every breed which I could purchase, or obtain, and have been most kindly favored with skins from several quarters of the world…. I have associated with several eminent fanciers, and have been permitted to join two of the London Pigeon Clubs.
Darwin states an excellent reason for his choice in the next sentence:
The diversity of the breeds is something astonishing. Compare the English carrier and the short-faced tumbler, and see the wonderful difference in their beaks…. The common tumbler has the singular and strictly inherited habit of flying at a great height in a compact flock, and tumbling in the air head over heels…. The Jacobin has the feathers so much reversed along the back of the neck that they form a hood…. The fantail has thirty or even forty tail-feathers, instead of twelve or fourteen, the normal number in all members of the great pigeon family; and these feathers are kept expanded, and are carried so erect that in good birds the head and tail touch.
These breeds are so different that any specialist, if “he were told that they were wild birds,” would assume major taxonomic distinction based upon substantial differences. “I do not believe,” Darwin writes, “that any ornithologist would place the English carrier, the short-faced tumbler, the runt, the barb, pouter, and fantail in the same genus.”
And yet, demonstrably by their interbreeding and their known history, all these pigeons belong to the same species, and therefore have a common evolutionary parent—the rock-pigeon,
Columba livia
. Darwin writes: “Great as the differences are between the breeds of pigeons, I am fully convinced that the common opinion of naturalists is correct, namely, that all have descended from the rock-pigeon (
Columba livia
).” (Darwin might have chosen the even more familiar example of dogs to make the same point, but he was not convinced that all dog breeds came from a common wolf source, whereas the evidence for a single progenitor of all pigeons seemed incontrovertible.) Only one step—the key analogy that powers the entire
Origin of Species
—remained to secure the most important argument in the history of biology, and to make pigeons the heroes of reform: If human breeding, in a few thousand years at most, could produce differences apparently as great as those separating genera, then why deny to a vastly more potent nature, working over millions of years, the power to construct the entire history of life by evolution? Why acknowledge the plain fact of evolution among pigeons, and then insist that all natural species, many no more different one from the other than pigeon breeds, were created by God in their permanent form?
Whitman, of course, did not disagree with Darwin’s focal contention
that
pigeons had evolved, but he strongly questioned Darwin’s opinion on
how
they and other species had arisen. Charles Otis Whitman (1842–1910), though scarcely a household name today, was the leading American biologist of his generation. He was, perhaps, the last great thinker to span the transition from the pre-Darwinian world to the rise of twentieth-century experimental traditions—for he had studied with Louis Agassiz, the last true creationist of real stature, and he lived to found and direct the symbol of rigorous modernism in American biology, the Marine Biological Laboratory at Woods Hole, still very much in vigorous operation. In his research, Whitman gained fame for meticulous work on
cell-lineage
studies in embryology—tracing the eventual fate of the first few embryonic cells in forming various parts of the body. In promoting this form of research as canonical in Woods Hole, and in establishing at his new laboratory the finest young American biologists, Whitman succeeded in bringing to this country the experimental and mechanistic traditions championed as the soul of modernism in Europe.
In this light, I have always had trouble remembering that Whitman’s main love in research lay in the opposite camp of “old-fashioned” and largely descriptive natural history—decades of work on the raising, breeding, and observation of pigeons. This passion even led to his death. In Chicago (where he served as professor of Biology at the University), on the first chill day of December, 1910, Whitman worked all afternoon in his backyard, hastily preparing winter quarters for his pigeons to save them from the cold. As a result, he developed pneumonia and died five days later. F. R. Lillie, once his assistant and then his successor at Woods Hole, eulogized his old boss: “In his zeal for his pigeons, he forgot himself.”
Unfortunately, Whitman died before completing and integrating his lifelong work on pigeons. He had published a few preliminary addresses (most notably, his offering in Saint Louis), but never the promised major statement. I can’t help thinking that the history of evolutionary thought might have been different had Whitman lived to promulgate and proselytize his non-Darwinian evolutionary ideas. His colleagues did gather his notes and data into a three-volume posthumous work on pigeons, finally published in 1919. But this work (the basis for my essay) was too disjointed, too incomplete, and, above all, too late to win its potential influence.
Darwin’s pigeon agenda extended beyond the simple demonstration of evolution. He also wished to promote his own theory about how evolution had occurred—natural selection. Again, he relied on argument by analogy: Pigeon breeds had been made by artificial selection based on human preferences for gaudiness (pouters, fantails) or utility (carriers, racers)—see figure. “When a bird presenting some conspicuous variation has been preserved, and its offspring have been selected, carefully matched, and again propagated, and so onwards during successive generations, the principle is so obvious that nothing more need be said about it.” (This quotation comes from Darwin’s most extensive discussion of pigeons—two long chapters in his 1868 book on
The Variation of Animals and Plants under Domestication
. Other statements in this essay are cited from the
Origin of Species
, 1859). But if selection is so undeniably the cause of small-scale evolution over millennia, why deny to nature the power for similar, but far greater, transformation over eons: “May not those naturalists who…admit that many of our domestic races have descended from the same parents—may they not learn a lesson of caution, when they deride the idea of species in a state of nature being lineal descendants of other species?”
Orthogeneticists like Whitman did not deny natural selection, but viewed Darwin’s force as too weak to accomplish anything beyond a bit of superficial tinkering. Natural selection, they argued, can make nothing and can only accept or reject the variation that arises naturally among differing organisms in an interbreeding population. If the genetic and embryological systems of organisms prescribe a definite direction to this variability, then natural selection cannot deflect the course of evolutionary change. Suppose, for example, that size in offspring only varied in a single direction from parental dimensions—that is, all kids ended up either the same size or taller than their folks. What could natural selection do? Darwin’s force could hasten an inherent trend by favoring the taller offspring. At most, selection could prevent a trend and keep the population stable, by eliminating the taller offspring and preserving only those of parental dimensions. But selection could not work counter to the inherent direction of variation because no raw material would be available for trends in any direction other than increasing size. Thus, selection would be a force subsidiary to an internal tendency for directional variation—or orthogenesis.
