Read Life's Ratchet: How Molecular Machines Extract Order from Chaos Online
Authors: Peter M. Hoffmann
Life's Ratchet: How Molecular Machines Extract Order from Chaos (33 page)

When Takeshi set up his experiments, it was already known that myosin V—unlike kinesin—took large 36-nm long steps. But there were still unanswered questions, such as how motion related to the use of chemical energy. To establish this link, Takeshi and his coworkers first primed their myosin V with a fluorescent label and supplied it with fluorescently labeled ATP. After focusing their microscope, they observed bright dots of different colors (they could not see the molecules directly, but only observed the fluorescent labels) moving along actin filaments in a choreographed step dance. First the ATP dot moved 18 nm, then the myosin dot moved 36 nm, and then the ATP dot moved another 18 nm, and then the cycle repeated (
Figure 7.7
). What did this mean? Takeshi and his colleagues could not distinguish between ATP, ADP-P (an ATP molecule in the process of being hydrolyzed—basically ADP with a nearby phosphate group), or ADP. All forms of the nucleotide shone with the same color and
brightness. Another complication arose when both heads were bound to a nucleotide. In this case, the researchers did not see two separate fluorescent dots, but just one dot, albeit one twice as bright. This was because the resolution of their microscope was not sufficient to distinguish two fluorescent labels so close to each other.
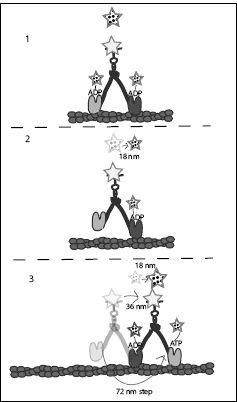
FIGURE 7.7.
The motion of myosin V and the associated fluorescent labels in Takeshi Sakamoto’s experiment. Fluorescent labels are shown as stars. Polka-dotted stars represent fluorescent labels attached to nucleotides or the visible fluorescent spot due to these labels (floating above each molecule). The size of the star indicates the brightness of the fluorescence. If both heads had nucleotides bound to them, one bright fluorescent spot was seen at the center of the myosin, while when only one nucleotide was bound to the myosin, a dimmer spot was seen at the location of the nucleotide bound head. The nondotted star represents the fluorescent label that is directly attached to the myosin molecule. Arrows indicate movement. See text for more explanation.
With this information at hand, and with the knowledge gained from previous experiments, Takeshi and his coworkers surmised that the first step in the cycle was the dissociation of ADP from the trailing head. Initially, when ADP was bound to both heads, the brightest location of the fluorescence was between the two heads. But as one ADP dissociated and blinked out, only the front head remained illuminated by its label. Thus the spot they saw moved from the center of the molecule to the leading head, while dimming to half its brightness. Since the heads were separated by 36 nm, this meant the spot moved by half this distance, or 18 nm.
Next, the fluorescent label attached to the myosin moved by 36 nm. This meant that following the release of ADP from the trailing head, this head made a 72-nm step, hand-over-hand, to propel the whole molecule
36 nm (remember, the other head was still attached and did not move). Almost simultaneously with this 36-nm motion, the ATP label moved 18 nm. Why 18 nm and not 36? Because in the meantime, ATP had attached itself to the forward-moving head, thus moving the center of the nucleotide fluorescence back to the center of the molecule.
It had previously been discovered that a myosin head, when bound to ATP or ADP-P, binds very weakly to the actin filament, although it binds strongly to actin in the presence of ADP or if the head is empty. Thus, very rapidly, prior to rebinding to the filament, the forward-moving head hydrolyzed its ATP into ADP-P. Then it released P and, now left with only ADP in its pocket, latched on to the actin filament. The timing of these steps, which happen very rapidly, is still not completely understood. Is phosphate released prior to actin binding, or after? Is it released before or after a conformational change of the head, which creates the forward tilt? These questions remain unanswered.
An even greater mystery surrounds the use of ATP hydrolysis in myosin V. For one, myosin V is completely different from kinesin, where ATP hydrolysis was used to detach the head from the filament. In the case of myosin, ATP hydrolysis apparently leads to the
attachment
of the head to the filament. But attaching a head to the filament does not cost energy; it should supply energy. So why supply even more energy at this step by hydrolyzing ATP? This does not make sense to me. If not for binding, how does myosin use the energy supplied by ATP hydrolysis? At the writing of this book, this is still a mystery.
When asked about this, Takeshi said his hunch was that the energy of ATP hydrolysis is not used directly to produce motion. If this is correct, the energy to provide motion comes from the energy of binding to the track, the release of internal strain in the molecule, and the thermal energy of the molecular storm. But then myosin would violate the second law. Thus, ATP hydrolysis needs to play the role of the reset step. For example, hydrolysis might prime a power stroke, the deformation that tilts the molecule forward to induce the trailing head to move in the forward direction. As the moving head swings forward, its long neck is oriented vertically. When ATP hydrolyzes, apparently just before binding to actin, the neck linker swings backward, releasing strain in the molecule. Upon binding, the neck linker swings forward again (probably powered by the binding
energy when the head binds to actin), providing the strain that tilts the molecule in the forward direction before it can take another step. So, again, the free energy supplied by ATP hydrolysis is used to reset the system. In the case of myosin, it may be used to temporarily release strain, to allow it to be applied again as soon as the molecule is in a position where this strain becomes useful to rectify its motion.
MYOSIN II: MUSCLE MOTION
Myosin V is no doubt fascinating due to its iconic, long-legged walking motion. But the original myosin, and the one most linked to the motions of animals that so fascinated biologists from Aristotle onward, is myosin II. The molecular machine that makes muscles work, myosin II was the first molecular motor discovered—by Andrew Huxley, in 1969. Strictly speaking, in evolutionary history, myosin II existed before muscles. “Nonmuscle” myosin II in cells pulls on actin filaments, which are part of the cell’s skeleton, and thereby helps the cells change their shape. As is common in evolution, once multicellular organisms evolved, the actin-pulling machines were co-opted to move the whole organism—using muscles.
It is quite easy to see how nonmuscle myosin may have turned into muscle myosin: Muscles are made of bundled bundles of muscle fibers— each fiber a single cell. Each cell contains myofibrils, which are structures composed of actin filaments interlaced with fibrous bundles of myosin II (
Figure 7.8
). Muscle cells are cells with an exaggerated network of actin and myosin—otherwise, myosin does what it has always done, even before muscles existed: pull on actin.
The structure of muscle and the presence of a molecule called myosin was known long before 1969, when Huxley proposed the first model to explain how an army of molecular motors could produce macroscopic motion. At the time, it was already known that muscle contraction was due to the relative sliding of two different filaments against each other: actin filaments and myosin filaments. But what generated the force that made the two filaments move relative to each other? Using electron microscopy of muscle fibers, Huxley suggested that there were distinct cross-bridges between the two filaments and that these bridges generated force and motion by tilting. The crossbridges turned out to be myosin heads,
and thus muscle motion was now attributed to the concerted efforts of legions of myosin molecules pulling on actin filaments.
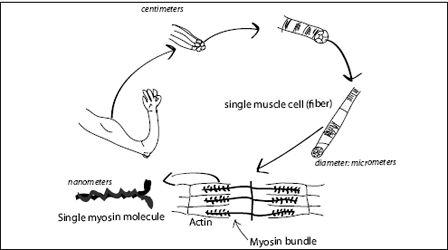
FIGURE 7.8.
The structure of muscle.
It is worth considering the scales involved in the various components of muscular movement. Muscles are macroscopic objects with a typical dimension of centimeters (or inches). As we zoom in, we first find that muscles are made of large strands, each of them a bundle of smaller strands, the muscle fibers. A single muscle fiber may be centimeters in length, but is only about 50 micrometers (a millionth of a meter or a 25,000th of an inch) in diameter. A muscle fiber is a very long cell. Inside the cells are bundles of actin and myosin. Each myosin head, the active force-generating part of the whole assembly, is only 30–40 nanometers in size. Every gram of muscle contains about 10
17
(100 million billion) myosin molecules.
The process by which myosin II generates motion is still not completely understood, but substantial progress has been made by structural (X-ray, electron microscopy), biochemical, fluorescence, and laser tweezer studies. The cycles of attachment and detachment, as well as ATP binding, hydrolysis, and release, are essentially the same for myosin II and myosin V. However, myosin II does not seem to be processive. It does not walk along actin filaments for long distances, as myosin V does. On the other hand, myosin II can produce faster motion than myosin V can.
As mentioned in
Chapter 6
, some people believe that myosin II is a weakly coupled motor, while others believe in a distinct, fixed-distance power stroke. Although myosin II was the first molecular motor discovered, it remains one of the most enigmatic. For example, as in myosin V, the role of ATP hydrolysis does not seem to be settled: In dozens of papers, most authors seem to reduce the function of ATP hydrolysis to an “increase in affinity to actin,” while a few researchers assume that ATP hydrolysis is associated with some prepower stroke. Moreover, as in myosin V, the ATP hydrolysis proceeds in several steps: splitting off a phosphate to create ADP-P, and only then releasing the phosphate, which results in a bound ADP. These steps happen too fast for scientists to clearly link them to mechanical motions of the myosin. Solving this problem will require the development of faster fluorescence techniques. The hydrolysis of ATP releases a fair amount of energy, so it is difficult to imagine that it would simply serve to make the myosin head more attractive for actin. Perhaps it plays a role in biasing diffusion, for example, by internally changing the strain in the molecule.
How, then, is muscle motion controlled? One explanation is that some mechanism controls the supply of ATP to muscle. If we want to move, our bodies supply ATP; if not, they shut off the ATP supply. But this is not how it happens. As a matter of fact, in myosin II, the binding of ATP allows the myosin head to detach from actin. Therefore, a lack of ATP leads to rigor mortis: The myosin gets stuck on the actin and the muscles harden up. Instead, the motion of muscle is controlled by a molecular switch called tropomyosin. Tropomyosin binds to actin and blocks the site where myosin takes hold of the actin filament. When we think about lifting an arm, nerves send a signal to the muscle cells, which release calcium ions. The calcium ions bind to the tropomyosin and it releases the actin, thereby exposing a patch to which myosin II can bind.
THE MYOSIN SUPERFAMILY
Myosin, like kinesin, forms a large family with many functions (
Figure 7.9
). Muscle myosin II generates power in muscles, while nonmuscle myosin II helps cells change shape, move, and divide. Myosin V is a short-distance transporter in our cells. What about the other myosin molecules? Myosin I, unlike myosin II and V, is a
single-headed
motor used in cell motion. It attaches to the cell membrane on one end and to actin at the other. Then it pushes actin filaments around as they are polymerized (generated). In this way, it shapes the internal skeleton of the cell and provides tension between the skeleton and the cell membrane.