Read What is Life?:How chemistry becomes biology Online
Authors: Addy Pross
What is Life?:How chemistry becomes biology (12 page)

Let us begin with a fundamental but unproven assumption, that life on earth was initiated from abiotic beginnings some period of
time after our solar system was formed some 4.6 billion years ago. That assumption forms the basis of the modern view which took shape in the 1920s through the joint contributions of the Russian biochemist, Alexander Oparin, and the influential British geneticist and evolutionary biologist, J. B. S. Haldane. An alternative scientific view, panspermia, invokes the idea that life originated from beyond the earth and was transported in some fashion to the prebiotic earth. That idea, proposed toward the beginning of the twentieth century by a well-known physical chemist, Svante Arrhenius, is, however, no longer seriously considered by the majority of researchers in the area, even though it has been supported by some well-known figures, including Francis Crick. A key difficulty with the panspermia proposal is that it does not really solve the problem of abiogenesis—the manner by which life emerged from inanimate beginnings—it merely transplants the problem to some other unidentified cosmic location. Regardless of its location, the question remains unchanged: how did life emerge from non-life?
Before addressing the origin of life question in greater detail, it is crucial to point out that the question has two quite distinct facets—
historical
and
ahistorical
, and only the
combined
insights of the two facets will be able to lead to a full and satisfying resolution of the problem. The historical aspect would seek to answer the
how
question—
how
did life emerge. That would involve deciphering the actual chemical events that transpired on the prebiotic earth—the particular chemical path followed, step by step, leading from inanimate materials through to simplest life. Key questions would
include: what were the molecular building blocks from which early life was constructed? What were the prevailing reaction conditions that enabled those building blocks to form, and once formed, what were the key intermediate steps along the long evolutionary road from those building blocks to simple life? As we will shortly see, not only has no broad agreement on these issues been reached, but practical knowledge of any kind regarding specific conditions on the prebiotic earth remains seriously wanting.
The ahistorical aspect would address the more general
why
question: why would inanimate matter of any kind, regardless of its structural identity, follow a pathway of complexification in the biological direction, eventually leading to some simple life form? I ask the question in the sense of identifying a driving force, seeking the same kind of insight that Newton sought when he asked ‘why do apples fall?’ Could the process, at least in principle, be induced in a range of different materials? What physicochemical principles could explain such an extraordinary chemical transformation? And with respect to this last question, can we go a step further and postulate the existence of a ‘physical’ driving force that would have directed inanimate matter to complexify in the biological direction? That question, as phrased, rests on an additional presumption, that the emergence of life was not a purely random event, but one that was induced by established physicochemical forces. We will discuss this presumption in greater detail subsequently. Thus the ahistorical perspective would not focus on the precise molecular identities of the relevant inanimate materials, but would seek out generalities—the
category
of material (or materials) that would likely have the propensity to become life, as well as the relevant physicochemical principles which would have induced
these materials to complexify into a simple life form. As we will see, here too the picture remains uncertain and highly controversial.
Given the paucity of information of any kind on the origin of life, ideally we would want to optimize our insights from both historical and ahistorical aspects in order to obtain as full a picture as possible. Before doing so, however, it will be useful to clarify the manner in which the two aspects interrelate. It turns out that each kind of information can serve as a means of obtaining information about the other kind. To illustrate how this interrelation works let us consider a simple physical analogy—a boulder that rolls down some mountain slope after being dislodged from its initial location (let us say due to water erosion or seismic activity), and finally comes to rest at the bottom of the slope. In this case ahistorical and historical aspects of that physical event are readily identified. The historical
how
question would be: from what initial location did the boulder begin its descent, and what trajectory did it follow? In principle there could be a large number of possible starting points, with each matched to a large number of potential trajectories. The second question—the ahistorical
why
question—would be to ascertain why, once dislodged, the boulder was induced to roll down the slope in the first place. Of course, in the case of a rolling boulder the answer to the second question is obvious—a gravitational force operates on all objects on the earth’s surface tending to lower their potential energy, so the physical reason for the boulder rolling down the slope is clear. Notice that the answer to the ‘why’ question is formulated in terms of a general law, independent of the specific location of the boulder, the nature of the terrain, etc.
There is an important message hidden within the rather trivial rolling boulder analogy. Historical knowledge and ahistorical
understanding impact on one another. For example, understanding the ahistorical aspect—the nature of the physical force responsible for boulder motion—would greatly assist in answering the historical question, that of boulder trajectory. Indeed, due to our knowledge of gravity we can safely exclude the possibility that the boulder simply levitated and floated through the air to its final location. Only boulder trajectories consistent with the action of a gravitational force would merit consideration. But the historical-ahistorical interplay also operates in reverse. Let’s assume for a moment that we are
not
familiar with the law of gravity. Obtaining information regarding the boulder’s trajectory would provide information as to the principles governing boulder motion in general. Uncovering the boulder’s trajectory would reveal that boulders apparently seek to lower their potential energy, i.e., they always roll from a higher location to a lower one—never in reverse, never uphill, and that the preferred trajectory is the pathway of steepest descent. So knowledge of a particular boulder’s trajectory, a historical event, would be a key step toward uncovering the rules governing boulder motion in general, the ahistorical aspect.
In the same way, if we want to address the historical origin of life question in its particulars, namely, to specify the starting materials and the particular set of reaction steps that led from those materials to early life forms, then knowing the general principles that govern the conversion of relatively simple molecular systems into the complex systems of life would be of considerable value. It would suggest the kinds of historical evidence we should be seeking. And vice versa, knowing the reactions that led to the conversion of inanimate matter into animate matter would greatly assist in uncovering the general principles that would have governed such a remarkable transformation.
But it is precisely at this point that we run into difficulty. Both kinds of information with respect to the origin of life problem are seriously lacking. Let me spell it out: since we are still struggling to understand the ahistorical principles, we don’t know which historical processes we should be seeking to uncover, and since we have no definitive historical evidence for the emergence of life at a particular location under particular conditions, we have no historical data to guide us in the elucidation and formulation of the relevant ahistorical principles. Catch 22!
So how to proceed? Before doing so, I will make a somewhat controversial statement with respect to the origin of life problem,
that the ahistorical question is the more significant one scientifically, and also the inherently more tractable one, the one that is less difficult to resolve.
As I will explain subsequently, this presumption will impact considerably on the nature of our discussion. I will describe how attempts to trace out plausible historical mechanisms for the emergence of life, without a prior understanding of the principles governing biological complexification, have not been able to resolve the problem, and may have even contributed to existing confusion. Often these hypotheses are untestable and, being highly specific in their formulation, do not address the more general ahistorical question. With this brief introduction, let us now examine the topic in some detail.
What historical information do we have regarding early life on the planet? On the basis of radiometric dating it is generally accepted that the earth was formed some 4.6 billion years ago, with the first 600–800 million years of the planet’s existence being thought to
have been too inhospitable for the emergence of life. During that initial period extensive bombardment from outer space would have been capable of evaporating the oceans and sterilizing the earth’s surface. The earliest historical evidence for the existence of life on earth is obtained from what is termed the palaeobiologic record—the microfossil remains of ancient microorganisms, and most recent findings date that early life at about 3.4 billion years old.
33
In any case all of these fossil findings point to relatively advanced cellular life, and therefore do not throw light on the earlier process of abiogenesis. Indirect evidence for the existence of earlier life going back 3.8 billion years ago is also claimed
34
though the issue remains controversial. In other words, the morphologically informative palaeobiologic record runs out after about 3.4 billion years when cellular life was already well established, so we must conclude that the palaeobiologic record in itself is unable to provide direct insights into the origin of life problem.
The second powerful technique for probing the history of life on earth is termed phylogenetic analysis, or, by its simpler name, sequence analysis. As the Nobel biophysicist, Max Delbrück, noted in a 1949 address before the Connecticut Academy of Arts and Sciences: ‘any living cell carries with it the experience of a billion years of experimentation’. In that spirit, sequence analysis enables us to construct a
tree of life,
which reveals the way all living things relate to one another by uncovering their evolutionary history. At the base of the tree we place the Last Universal Common Ancestor (LUCA)—the most recent living thing from which
all
life on earth proceeded to evolve—and from that base the trunk divides continually into more and more branches, each branch representing a new species. The very top of the tree represents all living species found on the earth today, while lower branches that end abruptly
represent species that became extinct. So start from any branch end, follow it back, and you will uncover the entire evolutionary record of that particular species.
How does sequence analysis reveal the structure of the tree of life? Two families of biomolecules that govern the nature and form of all living entities are nucleic acids and proteins, already mentioned in earlier chapters. Both of these groups of compounds involve long, chain-like molecules made up of monomeric units—nucleotides in the case of the nucleic acids, and amino acids in the case of the proteins. Since there are a variety of possible monomeric units from which the nucleic acid and protein molecules can be built up (4 nucleotide possibilities in the case of nucleic acids, 20 amino acid possibilities in the case of proteins), considerable variation in the
sequence
of the monomeric units within these two classes of biomolecules is possible. But here is the important point: the closer two species happen to be in evolutionary terms, the greater the similarity in the sequence of any biomolecule shared by the two species is likely to be. Carry out that comparative study for a large number of species and the genealogical relationship between different life forms, as expressed in a tree structure, can be established.
Sequence analysis in the 1970s started off with a spectacular result. Before those studies it had been assumed that
archaea
and
bacteria
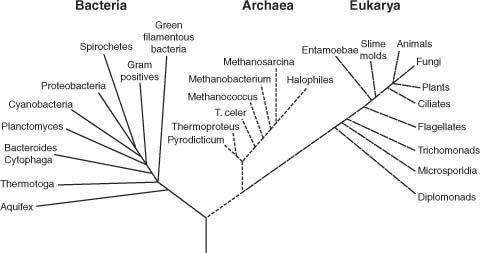
, two single-cell life forms, were closely related based on their similar morphologies and prokaryotic nature (primarily, the absence of a nucleus and other organelles). Archaea are commonly found in relatively harsh environments, such as hot springs and salt lakes, where traditional and more common bacterial life forms cannot survive. But in the late 1970s, primarily due to the pioneering sequence analysis work of Carl Woese, it was discovered that archaea are more closely
related to eukaryotic cells (those making up you and me) than to bacterial cells! As a result, the two seemingly closely related prokaryotic life forms were relegated to distinct and separate kingdoms. The tree of life, thought to be composed of
two
major kingdoms, Prokarya and Eukarya, was transformed into a tree with
three
kingdoms—Archaea, Bacteria, and Eukarya (illustrated in Fig. 4). Sequence analysis had proven to be a most powerful tool in elucidating genealogical relationship. A major step in constructing the tree of life had been taken. But that’s where the good news stops. Applying that powerful tool to the origin of life problem has proved disappointing. Sequence analysis has failed to throw more than minimal light on that problem. Let’s see why.